CARREGANDO OBJETO
Aguarde

O ciclo das pentoses fosfato é uma rota alternativa para a oxidação da glicose-6P, no citosol, sem gerar ATP.
Esta rota corresponde a um processo multicíclico onde:
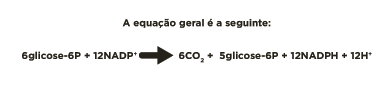
Clique aqui para ver a equação geral do ciclo das pentoses fosfato.
As células dos tecidos animais degradam a glicose 6-fosfato na via glicolítica (glicólise) até piruvato. Grande parte deste piruvato é oxidada a acetil-CoA, que por sua vez será oxidado no ciclo de Krebs (ciclo do ácido cítrico) formando ATP (principalmente, por fosforilação oxidativa).
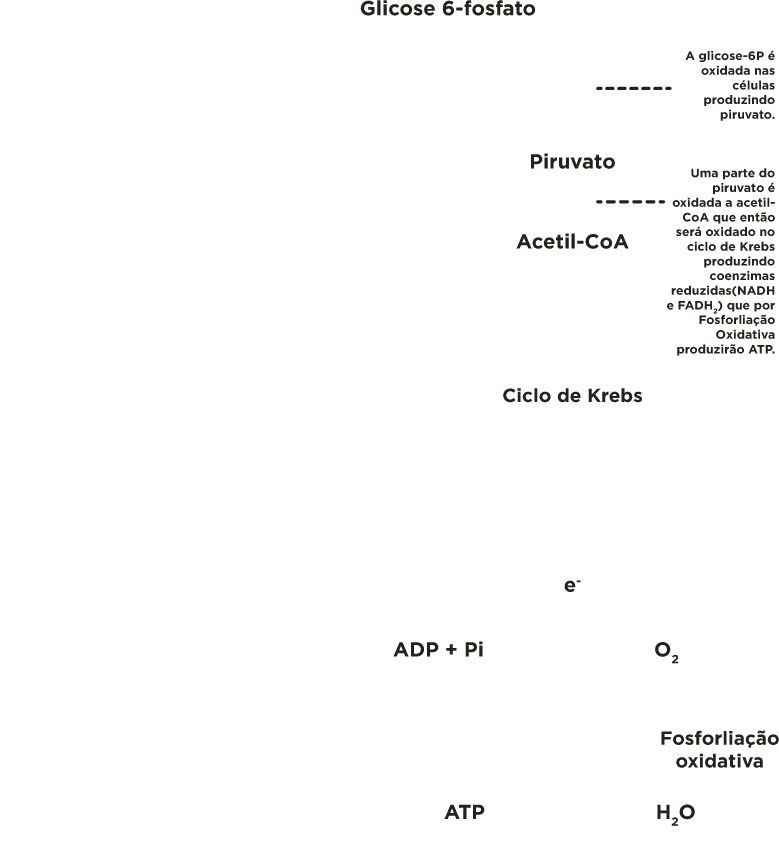
Porém, existem outros destinos catabólicos para a glicose 6-fosfato, entre eles: o ciclo das pentoses fosfato, também, denominado de via das pentoses fosfato ou via do 6-fosfo gliconato.

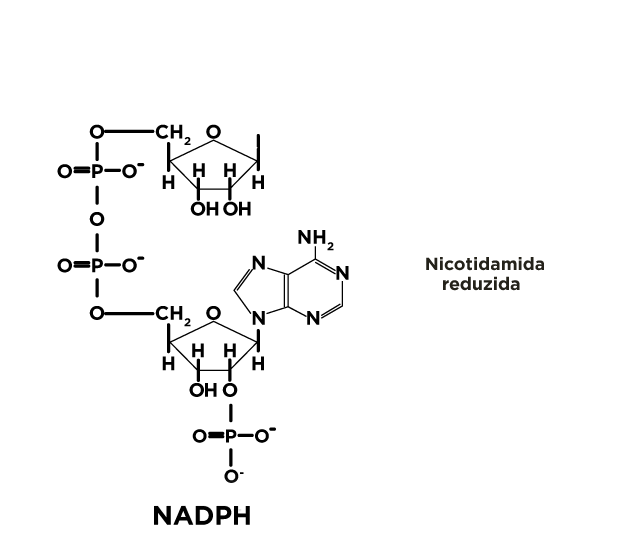
A via da pentoses fosfato é uma rota catabólica alternativa de oxidação da glicose-6P, que ocorre no citosol, sem produção de ATP, mas com geração de NADPH e pentoses fosfato.

O NADPH produzido pela via das pentoses fosfato é utilizado pelas células dos tecidos em que ocorre a síntese de grande quantidade de ácidos graxos (fígado, tecido adiposo, glândulas mamárias durante a lactação). Também ocorre onde há síntese de colesterol e hormônios esteróides (fígado, glândulas adrenais e gônadas).
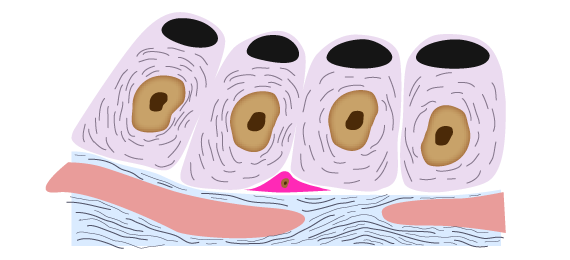
Os eritócitos, as células da córnea e do cristalino, também, possuem alta atividade da via das pentoses fosfato. Isto, para minimizar os efeitos deletérios das espécies reativas do oxigênio, pois estão diretamente expostos a ele. A manutenção do ambiente redutor (relação alta da concentração de NADPH para NADP+, assim como da glutationa reduzida para a oxidada) previne ou recupera o dano oxidativo sobre lipídios, proteínas e outras moléculas sensíveis.
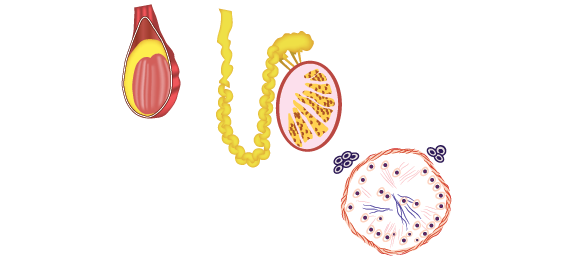
As células que se dividem rapidamente, como as da medula óssea, da pele, da mucosa intestinal, assim como as dos tumores, também, apresentam alta atividade da via das pentoses fosfato.
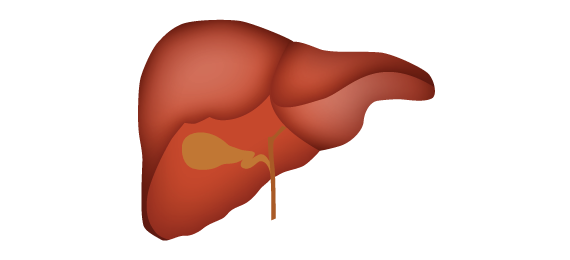
Biossíntese de ácidos graxos e do colesterol
e detoxificação hepática
Biossíntese de ácidos graxos e do colesterol
Biossíntese de ácidos graxos e do colesterol
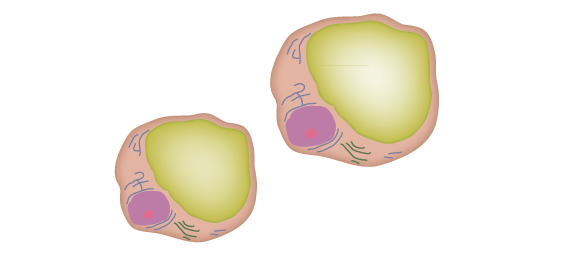
Biossíntese de hormônios esteróides
Biossíntese de hormônios esteróides
Biossíntese de hormônios esteróides
Manutenção do ambiente redutor através do NADPH

Manutenção do ambiente redutor através do NADPH
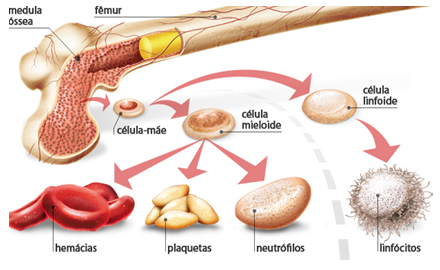
Células em rápida divisão celular
Células em rápida divisão celular
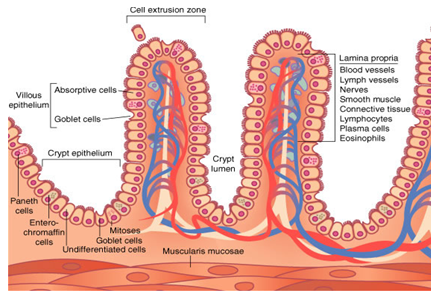
Células em rápida divisão celular
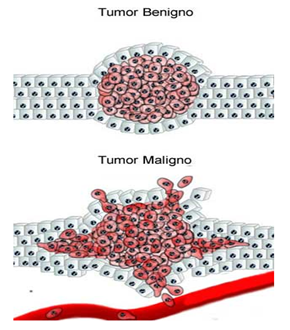
Células em rápida divisão celular
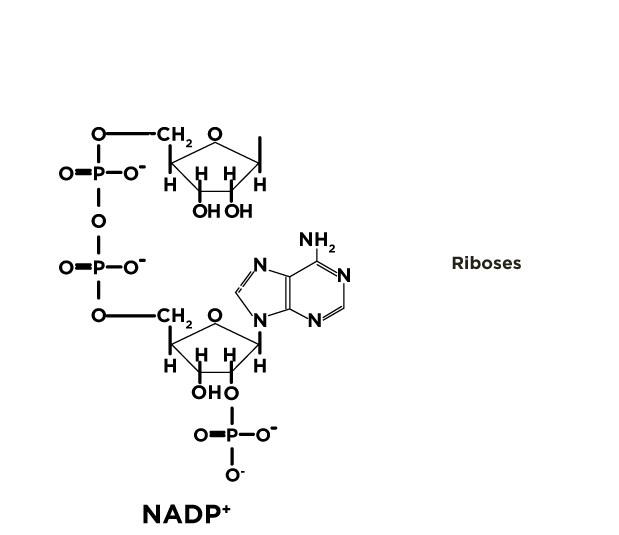
Espectro de absorção do NADP+ e do NADPH
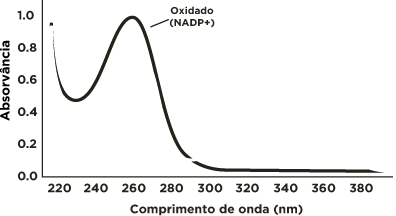
A coenzima oxidada absorve no comprimento de onda de 260 nm, enquanto que a coenzima reduzida(NAPDH) absorve tanto em 260nm quanto em 340nm
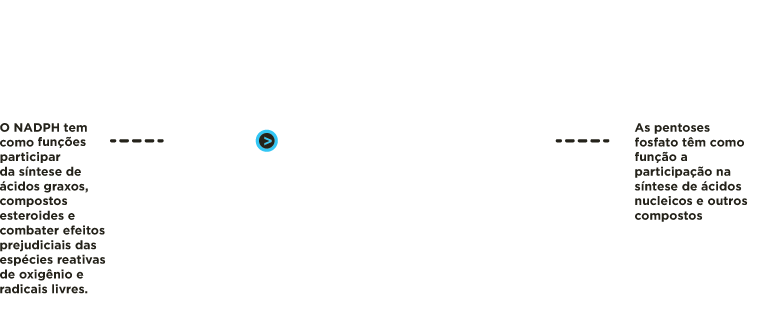
O NADPH é usado em diferentes biossínteses redutoras como a dos ácidos graxos, compostos esteróides e no combate a efeitos prejudiciais das espécies reativas de oxigênio.
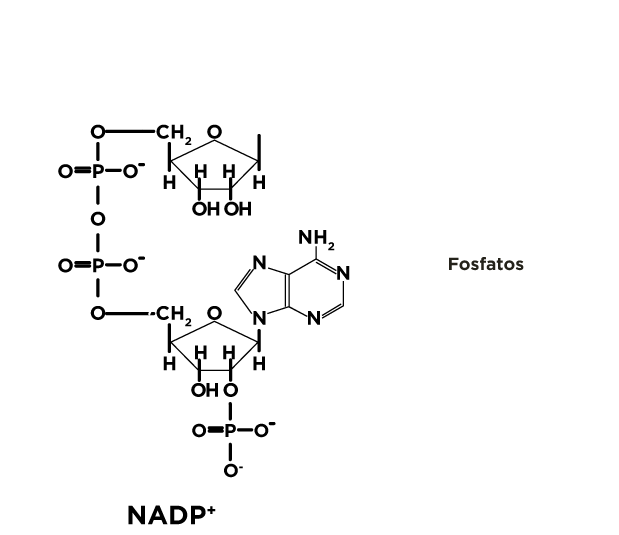
O outro produto essencial gerado na via das pentoses fosfato é a ribose 5-fosfato, que faz parte das estruturas químicas dos nucleotídeos (RNA, DNA, ATP) e coenzimas como NAD+/NADH, NADP+/NADPH, FAD/FADH2 e coenzima Q.
A via das pentoses fosfato é composta por duas fases: a oxidativa e não oxidativa.
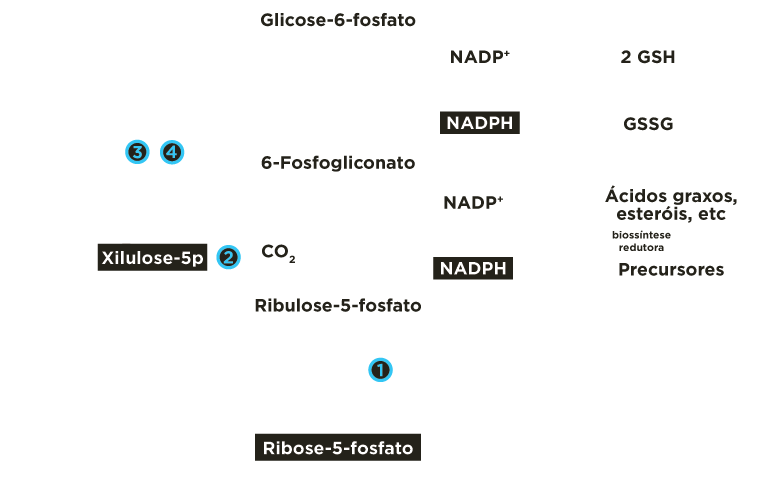
Na fase oxidativa ocorre a oxidação de 6 moléculas de glicoses 6-fosfato com formação de 6 moléculas NADPH, oxidação de 6 moléculas 6P-gliconato com formação de mais 6 moléculas NADPH e liberação de 6 moléculas dióxido de carbono e geração de 6 moléculas ribuloses 5-fosfato, que posteriormente se transformam em 6 ribose 5-fosfato.
Na fase não oxidativa, parte das ribuloses 5-fosfato continua a se isomerisar a ribose 5-P e parte se epimerisa a xilulose 5-P. Estas 2 pentoses fosfato reciclam e regeneram 5 moléculas glicoses 6-fosfato, permitindo a formação contínua de NADPH.
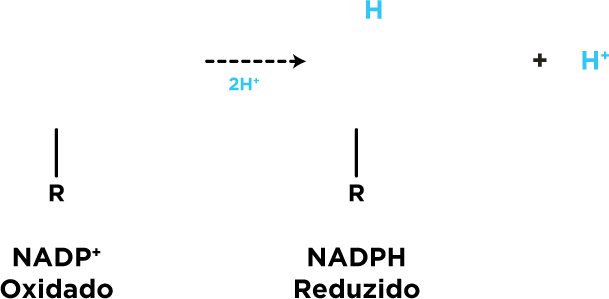
A glicose-6P é oxidada a 6P-gliconato com redução NADP+ . O NADPH tem como função combater efeitos prejudiciais das espécies reativas de oxigênio e radicais livres
. O NADPH tem como função combater efeitos prejudiciais das espécies reativas de oxigênio e radicais livres  , participar da síntese de ácidos graxos e compostos esteróides
, participar da síntese de ácidos graxos e compostos esteróides  .
.
A oxidação de 6P-gliconato com liberação de dióxido de carbono e geração de Ribuloses 5-fosfato e NADPH é a etapa seguinte  .
.
Posteriormente, a Ribulose-5-fosfato se transforma em ribose 5-fosfato 
Os NADPH formados na fase oxidativa são usados para produzir glutationa reduzida (GSH) a partir de glutationa oxidada (GSSG) e para participar das biossínteses redutoras
As riboses 5-fosfato são precursoras de nucleotídeos, coenzimas e ácidos nucleicos 
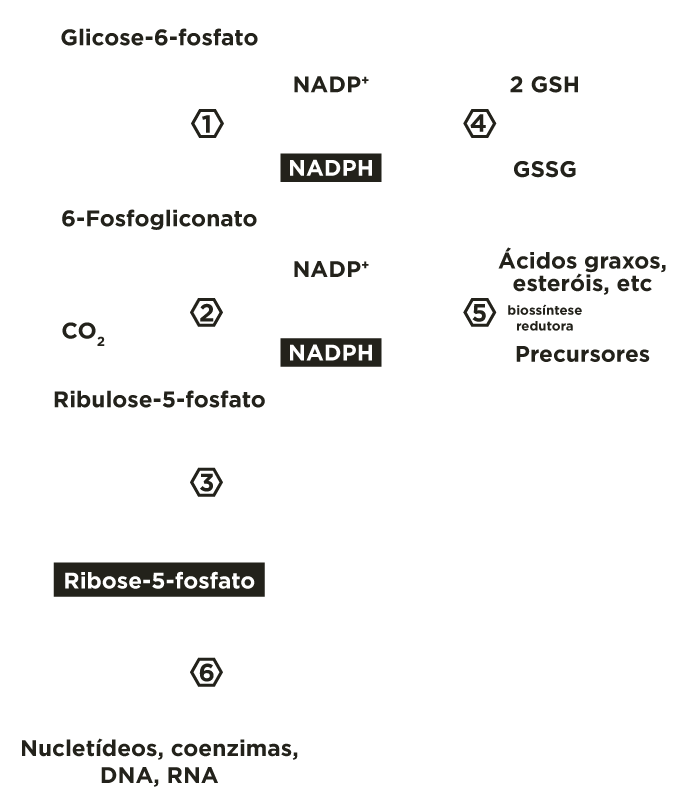
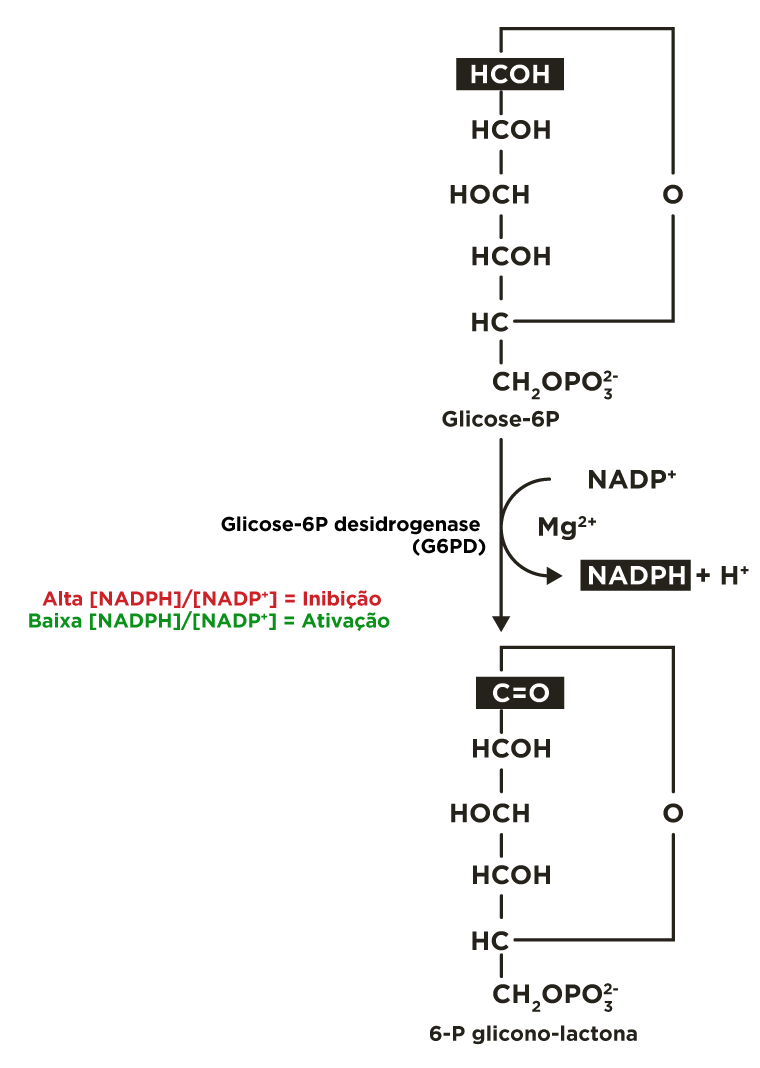
A via das pentoses fosfato inicia com a oxidação da glicose 6-fosfato pela glicose 6-P desidrogenase (G6PD) formando 6-fosfoglicono-lactona (éster intra-molecular) e NADPH. O equilíbrio desta reação está deslocado para o sentido de produção da coenzima reduzida.
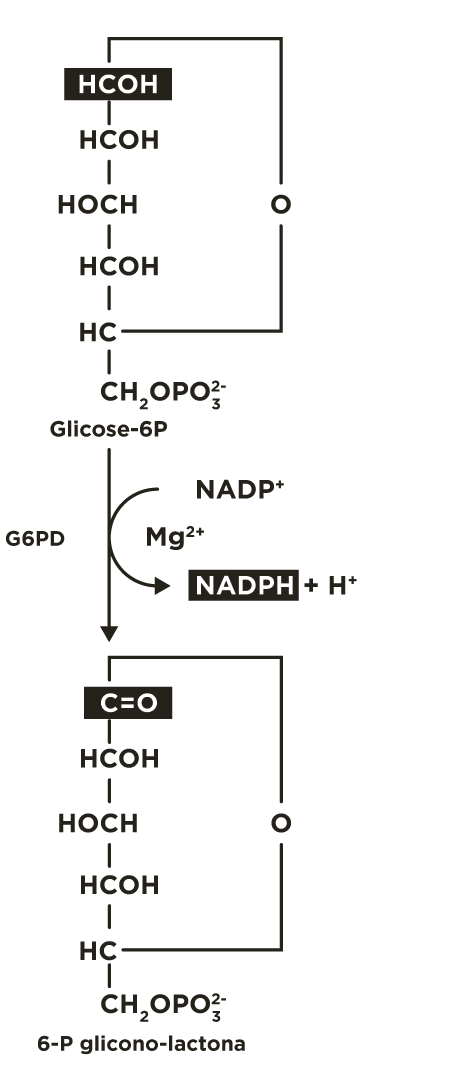
A 6-fosfoglicono-lactona é hidrolisada a 6-fosfogliconato por uma lactonase específica.

O 6-fosfogliconato é oxidado e descarboxilado pela 6-fosfogliconato desidrogenase (6PGD) liberando CO2 , formando a cetopentose ribulose 5-fosfato e outra molécula de NADPH.
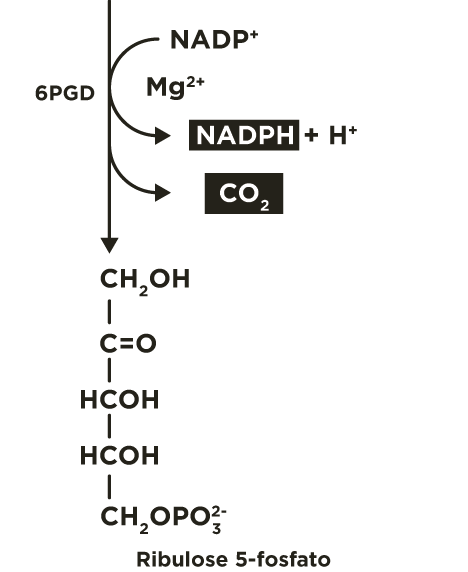
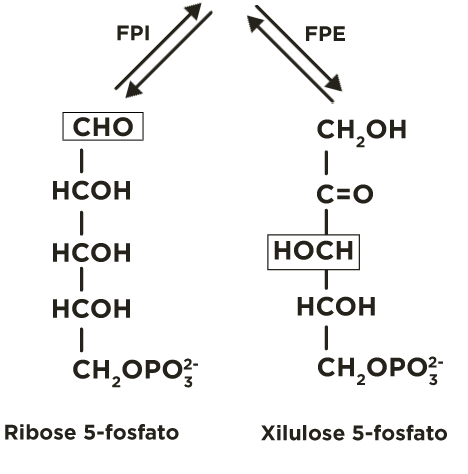
A ribulose 5-fosfato é convertida, principalmente, a seu isômero de função (aldose) ribose 5-fosfato, pela fosfopentose isomerase(FPI), e, secundariamente, a seu isômero de posição xilulose 5-fosfato, pela fosfopentose epimerase(FPE).
Em alguns tecidos a via das pentoses fosfato termina nesta etapa configurando o resultado de formação de NADPH, agente redutor, e principalmente riboses–fosfato, precursoras para a síntese de nucleotídeos.
Esta fase ocorre em tecidos que requerem, fundamentalmente, NADPH. Portanto, as ribuloses-5-fosfato, produzidas na fase oxidativa, são transformadas em ribose-5-fosfato por uma isomerase  ou em xilulose-5-fosfato por uma epimerase
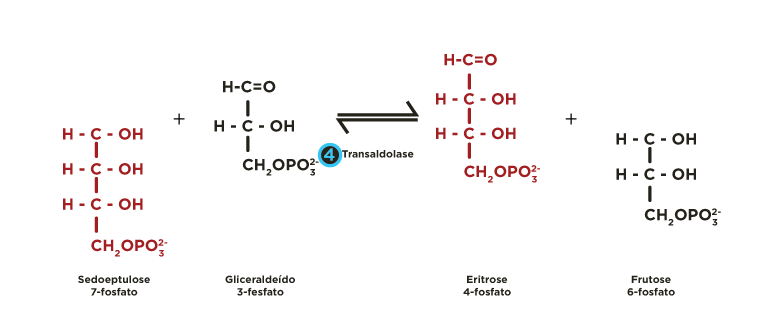
ou em xilulose-5-fosfato por uma epimerase  . Estas pentoses-5-fosfato são recicladas mediante a atividade de transcetolases
. Estas pentoses-5-fosfato são recicladas mediante a atividade de transcetolases  e transaldolases
e transaldolases  , regenerando glicoses 6-fosfato, que podem seguir novamente a fase oxidativa, permitindo a formação contínua de NADPH.
, regenerando glicoses 6-fosfato, que podem seguir novamente a fase oxidativa, permitindo a formação contínua de NADPH.
A ribulose 5-fosfato é transformada para ribose 5-fosfato por uma isomerase(FPI) .
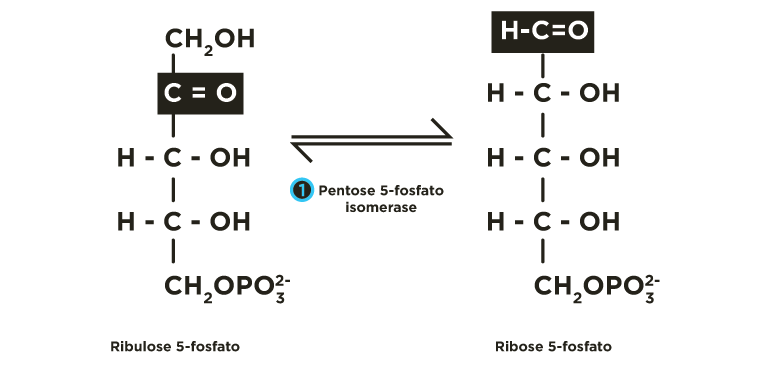
e para xilulose 5-fosfato por uma epimerase(FPE) .
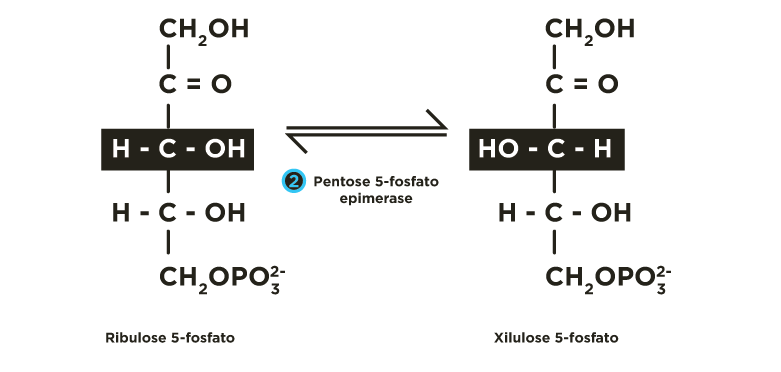
Ocorrem reações de transferências de fragmentos de 2 ou 3 carbonos catalisadas pelas enzimas transcetolase e transaldolase . Estes rearranjos moleculares geram intermediários da via glicolítica (gliceraldeído 3-fosfato, frutose 6-fosfato). A partir de 6 moléculas de glicose 6-fosfato são produzidas 6 moléculas de ribulose 5-fosfato que se reorganizam regenerando de 5 moléculas de glicose 6-fosfato.
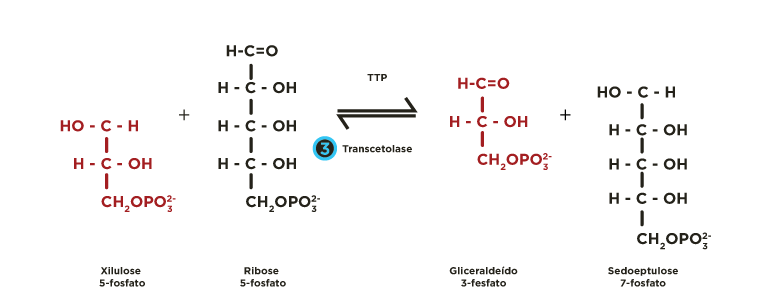
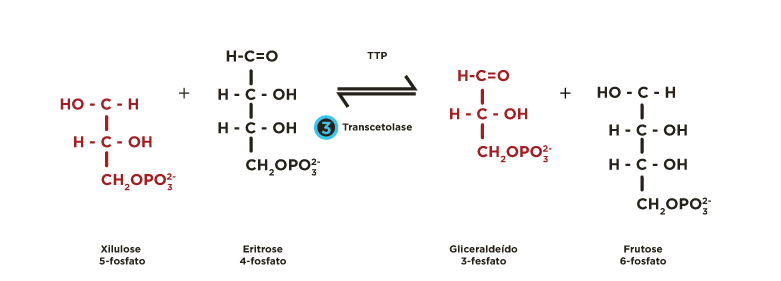






Rever animação.
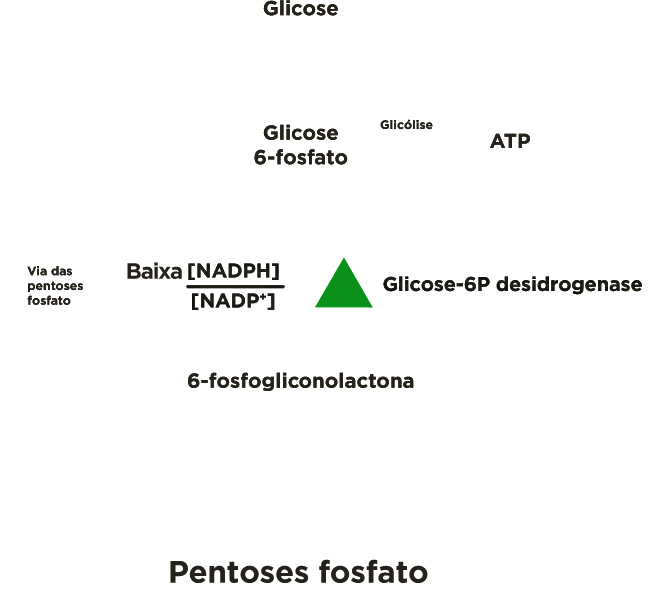
A entrada da glicose 6-P na via glicolítica ou na via das pentoses-P depende das necessidades momentâneas da célula, assim como, da relação entre as concentrações citosólicas de NADPH e NADP+ ([NADPH]/[NADP+]).
Quando o NAPDH é formado mais rapidamente do que é consumido nas reações de biossíntese em que participa, a sua concentração aumenta e, por conseguinte inibe alostèricamente a primeira enzima da via, a glicose 6-P desidrogenase, e, portanto mais glicose 6-P está disponível para a glicólise.
Por outro lado, quando o NAPDH é consumido mais rapidamente do que é produzido, a concentração de NADP+ aumenta, ativando alostèricamente a enzima glicose 6-P desidrogenase, aumentado o fluxo de glicose 6-P pela via das pentoses-P.
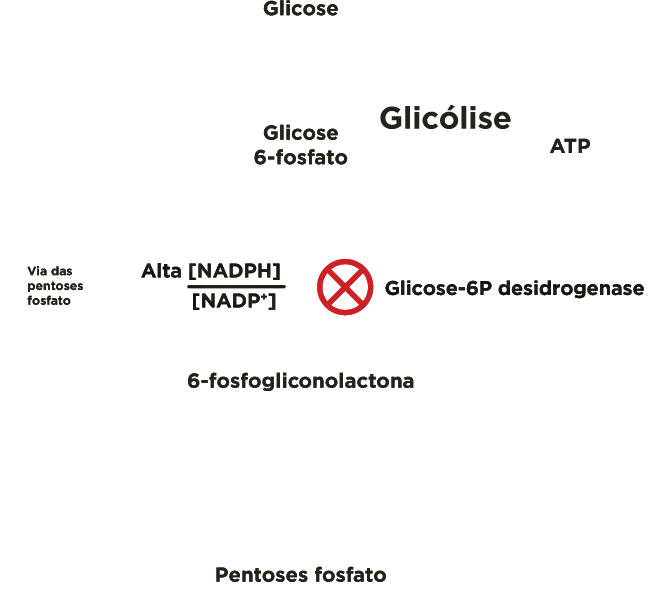
A enzima marca-passo da rota é a glicose-6P desidrogenase. Esta enzima é inibida quando a relação entre as concentrações de NADPH e NADP+([NADPH]/[NADP+]) estiver alta e é ativada quando a relação estiver baixa.
A entrada da glicose 6-P na via glicolítica ou na via das pentoses-P depende das necessidades momentâneas da célula, assim como, da relação entre as concentrações citosólicas de NADPH e NADP+ ([NADPH]/[NADP+]).
Quando o NAPDH é formado mais rapidamente do que é consumido nas reações de biossíntese em que participa, a sua concentração aumenta e, por conseguinte inibe alostèricamente a primeira enzima da via, a glicose 6-P desidrogenase, e, portanto mais glicose 6-P está disponível para a glicólise.
Por outro lado, quando o NAPDH é consumido mais rapidamente do que é produzido, a concentração de NADP+ aumenta, ativando alostéricamente a enzima glicose 6-P desidrogenase, determinando o aumento do fluxo de glicose 6-P pela via das pentoses-P.
Em breve, este objeto de aprendizagem receberá informações complementares como:
-Interações do ciclo das pentoses com a via glicolítica;
-Exemplos da participação de NADPH em várias reações químicas e processos biológicos: Biossíntese dos ácidos graxos, Sistema glutationa oxidada/glutationa reduzida, Sistema citocromo P450 monoxigenase, Reações da NADPH-Oxidase e da mieloperoxidase, Síntese de ácido nítrico;
-Fonte alternativa de NADPH;
-Patologias e vários processos biológicos relacionados: Deficiência da glicose-6-P desidrogenase, Papel do NADPH/glutationa na defesa celular contra os derivados altamente reativos do oxigênio.
NELSON, D. L.; COX, M.M. Princípios de Bioquímica de Lehninger. Tradução Fabiana Horn e colaboradores. Revisão técnica Carla Dalmaz e Sandra E. Farias. 6 ed. Artmed, Porto Alegre, 2014.
HARVEY, R.A.; FERRIER, D.R. Bioquímica Ilustrada. Tradução Carla Dalmaz e colaboradores. 5. ed. Artmed, Porto Alegre, 2012.
SMITH, C.; MARKS, A.D.; LIEBERMAN, M. Marks' Basic Medical Biochemistry – A clinical Approach. Second edition, Lippincott Wiliams & Wilkins, Baltimore, 2005.
BERG, J.M.; TYMOCZKO, J.L.; STRYER, L. Biochemistry. Fifth edition, W. H Freeman and Company, New York, 2002.
Profa. Vera Maria Treis Trindade
Profa. Chistianne Gazzana Salbego
Alisson Rocha
Design Gráfico
Gabriel da Cunha
Programação
Debora Schimitt
Assessoria pedagógica
Gabriela Perry
Coordenação NAPEAD
Marlise Santos
Assessoria pedagógica
Grupo de Criação de Objetos Educacionais em Bioquímica (GCOEB)
Profa. Dra Elena Aida Bernard
Colegas e alunos do Departamento de Bioquímica – ICBS - UFRGS