Embriologia Vegetal: Esporogênese e gametogênese nas plantas com flores
CARACTERIZAÇÃO DOS ESTRATOS PARIETAIS

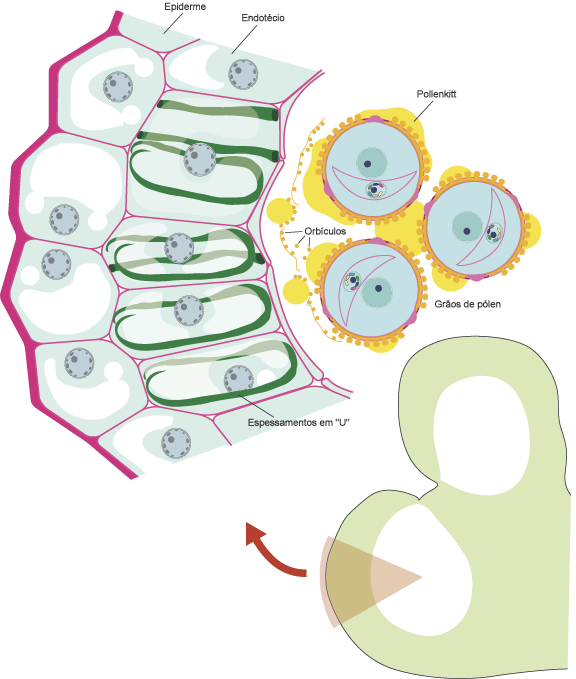
O desenho esquemático, acima, ilustra uma seção transversal da antera jovem. Nesta imagem, quatro estratos parietais podem ser identificados, do exterior para o interior: a epiderme, o endotécio, a camada média e o tapete.
A epiderme da antera é, tipicamente, fina na maturidade, podendo ocorrer o colapso, compressão, distenção ou ruptura de suas células quando, então, o endotécio constitui-se como a camada mais externa. Em alguns casos, a antera se reveste de tricomas epidérmicos. Em outros casos, a epiderme desenvolve faixas fibrosas (espessamentos de parede celular) a semelhança de um endotécio (sendo denominado exotécio) ou, ainda, a epiderme simula um tapete, com células binucleadas. Na abertura do microsporângio podem estar envolvidas, por exemplo, células epidérmicas, sem participação direta do endotécio. Durante a ontogenia da antera, as células do sítio de ruptura do estômio permanecem pequenas e com paredes delgadas. Já as fileiras de células epidérmicas de ambos os lados do sítio de ruptura diferenciam-se gradualmente em células estomiais, anticlinalmente alongadas e com cutícula, parede periclinal externa e parte das anticlinais espessadas. A ruptura do estômio coincide, aproximadamente, com o momento em que a flor se abre. As células estomiais se desidratam e, como resultado das forças de coesão-adesão entre as moléculas de água e as paredes celulares, se contraem, levando a abertura da antera.
O endotécio é a camada de células subepidérmicas que apresenta espessamentos parietais especializados, com função importante na deiscência (abertura) da antera. Os espessamentos parietais predominam nas paredes tangenciais internas e nas anticlinais. Entretanto, em alguns casos os espessamentos podem atingir as paredes tangenciais externas, resultando uma forma circular ou em anel, ou se apresentar sob forma helicoidal, ramificada, reticulada ou ainda como vários tipos de espessamento descontínuos. A ausência de espessamentos do endotécio é uma condição derivada e, muitas vezes, correlacionada à deiscência poricida (abertura da antera na forma de um póro apical), apesar de algumas famílias de Angiospermas apresentarem esse tipo de deiscência combinada com a presença de um endotécio espessado.
A constituição química dos espessamentos da parede das células do endotécio é, em geral, celulósica; em outros casos acrescido de pequena quantidade de material péctico e lignina. Geralmente está limitado a parte protuberante do esporângio, porém o conectivo também pode desenvolver, em uma ou mais camadas de células, esse tipo de espessamento.
A camada média pode ser formada ou não durante a ontogênese da parede do esporângio dependendo dos estratos parietais diferenciados. Quando presente o número de camadas é variável, podendo ocorrer 1 a 2 camadas de células, excepcionalmente até 5 estratos. Em geral são camadas transitórias ou efêmeras, sendo obliteradas ao final do desenvolvimento. Em algumas espécies são persistentes até a deiscência da antera. Em alguns casos, a camada média pode desenvolver espessamentos fibrosos semelhantes ao endotécio.
O tapete é camada mais interna dos estratos parietais e está constituído, em geral, por uma camada de células, podendo ocorrer bisseriado, multisseriado ou com uma camada de células na face voltada para a epiderme e multisseriado na face voltada para o conectivo. Devido à proximidade ao tecido esporogênico, o tapete apresenta as seguintes funções: nutrição do tecido esporogênico (arquespório) e dos andrósporos; secreção de calase durante a separação das tétrades ao final da esporogênese; síntese de esporopolenina para a formação da parede da parede dos grãos de pólen (esporoderme); produção dos orbículos (corpúsculos de Ubisch ou partículas de esporopolenina); síntese e liberação de materiais, como “pollenkitt” (material lipídico, flavonóides, carotenóides e produtos da degradação de proteínas do tapete), trifino (termo coletivo para “cobertura do pólen”, constituído de uma mistura de substâncias hidrofóbicas derivadas da dissolução do tapete, aparentemente formado por material protéico), enzimas, proteínas de reconhecimento e outras substâncias, sobre o grão de pólen; finalmente, pode ser responsável, em alguns casos, pela esterilidade masculina, citoplasmática e genética. Nas anteras jovens o tapete mantém a continuidade com as células-mãe de andrósporos através de plasmodesmos, pelos quais os nutrientes chegam a essas células, até a formação dos meiócitos. Essas conecções são interrompidas quando o espessamento de calose é depositado na parede interna das células-mãe de andrósporos durante o inicio do processo meiótico. As células do tapete são inicialmente uninucleadas, porém, na maioria das plantas, divisões mitóticas dos seus núcleos, não acompanhadas de citocinese, proporcionam o surgimento de células 2-, 4-, ou, algumas vezes, 16-nucleadas. Frequentemente, através de fusões nucleares, as células do tapete se tornam poliplóides.
O tapete pode ser classificado em dois tipos básicos: secretor ou glandular, quando o tapete permanece sempre adjacente aos demais estratos parietais, circundando o lóculo do esporângio, ou ameboidal ou periplasmodial, quando ocorre a protrusão das células para o interior do lóculo, com conseqüente fusão dos protoplastos, formando um plasmódio cenocítico. Em geral, as células do tapete perdem suas paredes celulares periclinais internas e radiais, no tapete secretor ou todas as paredes, no tapete ameboidal. Este evento ocorre no fim da meiose e formação das tétrades de micrósporos. Isto parece ter grande importância funcional para os processos de secreção de substâncias para o interior do lóculo.
Orbículos (ou corpúsculos de Übisch) são partículas de forma e tamanho variado que revestem a superfície interna das células do tapete secretor. Sua síntese ocorre no citoplasma das células do tapete. Para seu processo de polimerização e transporte, estão envolvidas vesículas associadas ao Golgi (dictiossomos) e retículo endoplasmático, as quais são secretadas através da membrana plasmática na forma de pro-orbículos. Ao contato com o fluido locular, essas vesículas rapidamente são impregnadas por esporopolenina, transformando-se em orbículos. Em estágios mais avançados da esporogênese, e em fases subseqüentes, o conjunto de orbículos forma a membrana tapetal (junto ao lóculo). Adicionalmente, a síntese de esporopolenina junto às paredes tangenciais externas do tapete, adjacentes à camada média, forma a membrana peritapetal. Com a degeneração do tapete, a membrana tapetal e peritapetal formam um “saco” envolvendo os grãos de pólen maduros e o conteúdo locular da antera.
A figura abaixo ilustra, esquematicamente, uma seção transversal de uma antera madura, em uma flor deiscente (aberta). Podem ser vistos os orbículos e o pollenkitt no interior da cavidade locular. Note que os únicos estratos parietais restantes são a epiderme e o endotécio.